←На предыдущую страницу
Кровообращение хордовых
Цели:
В ходе эволюции многоклеточных животных кровеносная система сформировалась на месте рудиментов первичной полости тела, вытесненной у высших животных вторичной полостью тела, или целомом. Кровеносная система приняла на себя в организме высших животных универсальную транспортную роль передвижение питательных веществ, усвоенных органами пищеварения, и кислорода от органов дыхания ко всем органам тела, перенос продуктов метаболизма к органам выделения, углекислоты — к органам дыхания, доставка гормонов от эндокринных желез к органам, где данные гормоны будут действовать. В силу этого кровеносная система стала и одной из важнейших интегрирующих систем организма, обеспечивающих его целостность. Кровеносная система имеется у немертин кольчатых червей, моллюсков, членистоногих. иглокожих, полухордовых и хордовых. Ее строение у животных разных типов сильно варьирует. Два основных продольных сосуда — спинной и брюшной, тянущиеся соответственно над и под кишечником, связаны друг с другом поперечными сосудами; последние кольцеобразно охватывают кишечник и дают ответвления к покровам и различным внутренним органам. У животных всех типов, условно объединяемых названием «беспозвоночные» кровь по брюшному сосуду течет назад, а по спинному вперед. У хордовых наоборот: по брюшному сосуду кровь течет вперед, а по спинному назад. Пульсирующей функцией, обеспечивающей движение крови, у хордовых обладает брюшной сосуд или его производное — сердце, у беспозвоночных эту функцию несет спинной сосуд. У общих предков вторичноротых животных (к ним принадлежат полухордовые, иглокожие и хордовые) кровеносная система имела очень примитивное строение, представляя собой систему лакун, т. е. полостей без эпителизированных стенок, где направление кровотока еще не установилось. В дальнейшем совершенствование кровеносной системы происходило независимо и по-разному в различных ветвях ствола вторичноротых. Поэтому состояние кровеносной системы хордовых сформировалось особым путем и не может быть выведено из развитой кровеносной системы беспозвоночных — как вторичноротых, так и (тем более) первичноротых, к которым принадлежат кольчатые черви, членистоногие, моллюски и др.
Среди хордовых наиболее примитивной кровеносной системой обладают животные подтипа бесчерепных.
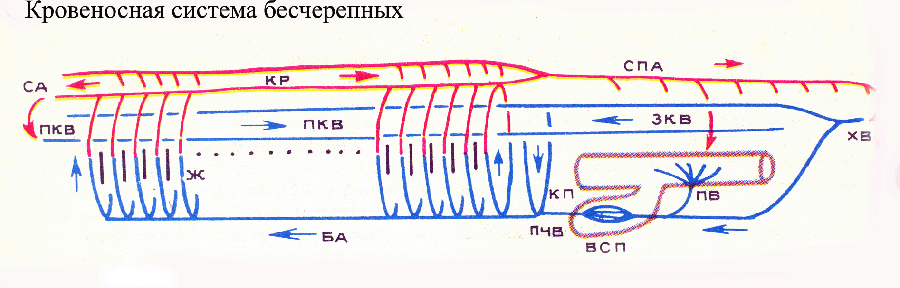
У типичного представителя бесчерепных — ланцетника — под глоткой тянется продольный сосуд — брюшная аорта, играющая роль основного пульсирующего органа. По брюшной аорте кровь течет вперед. От этого магистрального сосуда начинаются многочисленные (около 100 пар) жаберные артерии, которые расположены в перегородках между жаберными щелями, пронизывающими боковые стенки глотки. Основания жаберных артерий пульсируют, давая добавочный импульс току крови. Через жаберные щели непрерывно процеживается вода, и здесь происходит газообмен между кровью и водой: кровь в жаберных артериях обогащается кислородом и отдает углекислый газ, т. е. становится артериальной. Обогащенную кислородом кровь жаберные артерии доставляют на спинную сторону глотки, где они впадают в пару продольных сосудов — корни спинной аорты. Впереди корни аорты переходят в сонные артерии, несущие арте кровь в головной конец тела, корни аорты впадают в продольный спинной сосуд — спинную аорту, по которой артериальная кровь течет назад. От спинной аорты отходят артерии к различным внутренним органам и к кожным покровам, где артерии распадаются на капилляры. Из хвостовой области тела кровь собирается в хвостовую вену, впереди впадающую в подкишечную вену. Последняя собирает кровь от кишечника, а затем в печеночном выросте кишки вновь распадается на капилляры, образующие воротную систему печени. Из нее печеночная вена несет кровь в брюшную аорту. У ланцетника имеются еще две пары магистральных вен: передние и задние кардинальные, тянущиеся от переднего и заднего концов тела к средней его части, где они впадают в пару коротких Кювьеровых протоков. Последние несут кровь в задний конец брюшной аорты. Таким образом, брюшная аорта получает обедненную кислородом (венозную) кровь.
У второго подтипа хордовых — оболочников — кровеносная система подверглась редукции. Как и целый ряд других органов, в связи с переходом к неподвижному (прикрепленному) образу жизни на морском дне. Освоение такого образа жизни в эволюции обычно ведет к морфо-физиологическому регрессу. У типичных представителей оболочников асцидий — под желудком располагается трубкообразное сердце, от которого отходят два магистральных сосуда: спереди — жаберная артерия, тянущаяся по брюшной стороне глотки и отдающая ветки к перегородкам между жаберными отверстиями, позади — кишечная артерия, которая разветвляется на сосуды, идущие к внутренним органам. В конечном итоге артерии впадают в лакуны, т. е. кровеносная система оболочников не замкнута. Одна из таких лакун — жаберно-кишечный синус — лежит над глоткой и собирает кровь из жаберной области. Интересно, что направление пульсации сердца у оболочников периодически меняется: сердце попеременно гонит кровь то вперед, в жаберную артерию, то назад, в кишечную. Некоторые ученые видят в этом примитивную черту — еще не установившееся направление кровотока. Уникальной особенностью оболочников является их кровяной пигмент — гемованадин, в котором вместо ионов железа (как в гемоглобине остальных хордовых) или меди (как в гемоцианине беспозвоночных ряда групп) используются ионы ванадия. Эволюция третьего подтипа хордовых — позвоночных — в целом шла в направлении морфо-физиологического прогресса, что, в частности, ярко проявилось в последовательном совершенствовании кровеносной системы: ее дифференциации, интеграции, интенсификации функций в ходе филогенеза позвоночных. У всех позвоночных центральным пропульсаторным органом является сердце. У миног, представителей наиболее примитивного класса современных позвоночных — круглоротых, сердце имеет три отдела (камеры):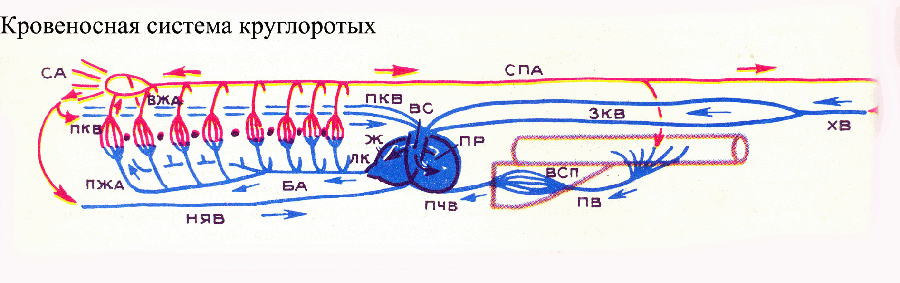
венозный синус (венозная пазуха), предсердие и желудочек . От желудочка отходит брюшная аорта, начало которой расширено (луковица аорты). Впереди брюшная аорта разделяется на пару поджаберных артерий. От брюшной аорты и поджаберных артерий начинаются 8 пар приносящих жаберных артерий, распадающихся в жабрах на капилляры (этого не было у бесчерепных). От жабер обогащенная кислородом кровь поступает в выносящие жаберные артерии, которые впадают в спинную аорту. В целом схема кровеносной системы круглоротых сходна с таковой бесчерепных, но отличается наличием еще одной магистральной вены (нижней яремной), выносящей кровь из вентральной части головы, и отсутствием Кювьеровых протоков (специфическая черта круглоротых). Обе пары кардинальных вен, печеночная и нижняя яремная вены впадают непосредственно в венозную пазуху. Еще одной специфической особенностью кровеносной системы круглоротых, является ее неполная замкнутость: жаберные мешки окружены околожаберными синусами, заполненными кровью и представляющими собой кровеносные лакуны. В связи с этим активное участие в циркуляции крови принимают мышцы жаберного аппарата, сокращение которых выталкивает кровь из лакун и сосудов жаберной области. У другой группы круглоротых — миксин — работе сердца помогают также пульсирующие участки ряда вен (кардинальных, хвостовой и воротной вены печени).
У всех остальных позвоночных — челюсторотых — по сравнению с круглоротыми был достигнут существенный прогресс в организации кровеносной системы. Она стала полностью замкнутой, сердце полностью приняло на себя роль центрального пульсирующего органа. Это сделало возможным повышение кровяного давления и интенсификацию кровотока, что способствовало общему повышению уровня жизнедеятельности. У хрящевых рыб (акулы, скаты, химеры) сердце состоит из 4 камер:

к З камерам, известным нам по круглоротым, добавился еще артериальный конус. Функция его — распределение крови между передними и задними приносящими жаберными артериями. Парные поджаберные сосуды, имевшиеся у круглоротых, редуцированы. У большинства хрящевых рыб жабры сохраняются лишь на 4 жаберных дугах и на подъязычной дуге висцерального аппарата; соответственно, имеется 5 пар приносящих жаберных артерий. Как и у бесчерепных, магистральные вены впадают не прямо в сердце, а в пару Кювьеровых протоков. В отличие от круглоротых, у хрящевых рыб нижняя яремная вена парная, а на пути крови из хвостовой области имеется еще одна капиллярная сеть — в почках (воротная система почек).
Класс костных рыб объединяет ряд филогенетически рано обособившихся друг от друга групп, у которых кровеносные системы существенно различаются по строению. У наиболее распространенных и много численных костных рыб (к ним относится большинство наших обычных пресноводных рыб подвергся редукции артериальный конус сердца, и оно стало трехкамерным, как у круглоротых. В начале брюшной аорты имеется расширение (луковица аорты). Развиты лишь 4 пары приносящих жаберных артерий. Воротная система левой почки редуцирована. Наземные позвоночные возникли от древних кистеперых рыб, обитавших в пресных мелководных, богатых растительностью тропических водоемах, отчасти заболоченных. В таких водоемах часто возникал дефицит растворенного в воде кислорода (в связи с усиленным гниением отмиравших растений). Важным приспособлением к жизни в этих условиях было развитие легких, позволявших при дефиците кислорода в воде дышать атмосферным воздухом. Среди современных костных рыб близкий образ жизни ведут родственные кистеперым рыбам двоякодышащие, тогда как единственный доживший до наших дней вид кистеперых — латимерия — обитает в морских глубинах и легкие для дыхания не использует. Морфо-физиологической особенностью рыб, дышащих и жабрами, и легкими (древние кистеперые и современные двоякодышащие), является решение проблемы рационального использования обоих органов газообмена. Дело в том, что при нормальном для рыб жаберном дыхании обогащенная кислородом кровь из передних жабер поступает в голову, тогда как из задних — в спинную аорту, несущую кровь назад по всему телу. Легкие сформировались позади глотки, т. е. позади самых задних жабер. И поэтому если кровь из легких, как из жабер, направить прямо в корни аорты, то эта артериальная кровь поступит в спинную аорту, а органы головы (и, в частности, головной мозг, особенно н уждающийся в постоянном поступлении кислорода) не смогут ее использовать. Поэтому возникает необходимость нового перераспределения крови. Эта проблема была решена возвращением артериальной крови из легких в сердце (т. е. образованием дополнительного легочного, или малого, круга кровообращения) и формированием в сердце особых приспособлений для перераспределения крови.
У двоякодышащих
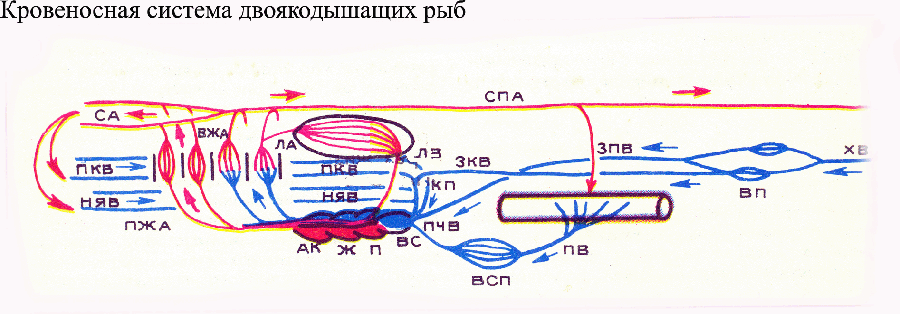
сердце состоит из венозного синуса (получающего, как обычно у рыб, венозную кровь), предсердия, желудочка и артериального конуса. Венозный синус впадает в правую половину предсердия, тогда как в левую его половину впадают легочные вены, приносящие артериальную кровь из легких. В предсердии, желудочке и артериальном конусе имеются неполные внутренние перегородки (в частности, спиральный клапан артериального конуса), позволяющие разделить кровь, поступившую из левой и правой половин предсердия. Из левой половины предсердия артериальная кровь поступает в нижнюю часть сильно укороченной брюшной аорты и по ней — к двум передним парам приносящих жаберных артерий; затем эта кровь поступает по сонным артериям в голову. Из правой половины предсердия венозная кровь направляется в верхнюю часть брюшной аорты и по ней — к двум задним (ближайшим к сердцу) парам жаберных артерий. Подчеркнем, что легочные артерии начинаются от четвертой (самой задней) пары выносящих жаберных артерий. Механизм распределения крови в сердце двоякодышащих рыб лабилен и позволяет направлять больше или меньше крови в передние или задние жаберные артерии в зависимости от физиологического состояния рыбы и от того, какой орган (легкие или жабры) в данный момент работают интенсивнее. В венозной системе у двоякодышащих рыб редуцирована правая задняя полая вена, зато появился новый магистральный венозный ствол — задняя полая вена, несущая кровь из воротной системы почек прямо в венозный синус, минуя Кювьеровы протоки. У наземных позвоночных (тетрапод) произошли значительные изменения в строении кровеносной системы по сравнению с рыбами (хотя общее направление этих перестроек намечено уже у двоякодышащих). У всех тетрапод брюшная аорта отчасти редуцирована, отчасти разделена на несколько обособленных артериальных стволов, начинающихся от сердца, причем это разделение произошло по-разному у предков современных земноводных, пресмыкающихся и млекопитающих. У современных земноводных
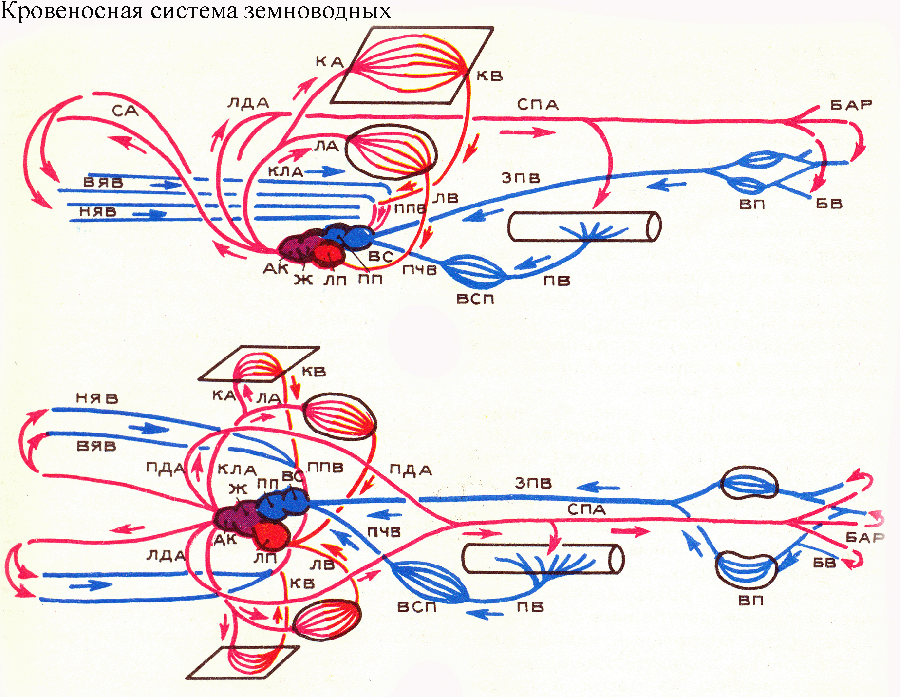
от артериального конуса сердца начинаются три пары сосудов: сонные артерии, дуги аорты (которые на спинной стороне сливаются, образуя спинную аорту) и легочные (а у бесхвостых амфибий — кожно-легочные) артерии. Первые из них соответствуют 1 паре жаберных артерий с собственно сонными артериями рыб; вторые — II паре жаберных артерий с корнями аорты рыб; третьи — IV паре жаберных артерий с легочными артериями рыб (у хвостатых амфибий иногда сохраняются две пары дуг аорты, соответствующие II и III парам жаберных артерий). У рыб все эти сосуды связаны друг с другом над глоткой корнями аорты; такая связь сохраняется и у некоторых тетрапод в виде сонных протоков между сонными артериями и дугами аорты, а также Боталловых протоков между дугами аорты и легочными артериями. По сонным артериям кровь направляется к голове; по дугам аорты — в спинную аорту и по ней к различным органам тела; легочные (или кожно-легочные) артерии обеспечивают кровоснабжение органов дыхания. У земноводных несовершенство механизма вентиляции легких не позволяет этим животным ограничиться легочным дыханием, и кожа у них является важным дополнительным органом дыхания. У хвостатых и безногих амфибий, легочные артерии не объединены с кожными. Кровоснабжение кожных покровов, как и у других позвоночных, осуществляется ветвями спинной аорты.
Венозная система земноводных отличается от таковой рыб редукцией задних кардинальных вен, которые замещаются задней полой веной, получающей кровь из воротных систем почек и печени и доставляющей ее в венозную пазуху сердца. Передние кардинальные вены у наземных позвоночных сохраняются, но получают новое название: их традиционно именуют верхними яремными венами. Кювьеровы протоки у тетрапод называют передними полыми венами. Сердце земноводных состоит из пяти отделов: венозного синуса (пазухи), левого и правого предсердий, желудочка и артериального конуса. В венозный синус впадают передние и задняя полые вены. В левое предсердие впадают легочные вены, приносящие артериальную кровь из легких. В желудочке кровь из левого и правого предсердий в определенной степени смешивается. Нередко это упрощенно объясняют примитивностью и несовершенством сердца земноводных, но в действительности дело обстоит сложнее и интереснее. Объединение в желудочке сердца земноводных крови, поступающей из левого и правого предсердий, имеет важное функциональное значение: если бы такого смешивания крови не было, земноводным не удалось бы эффективно использовать кожное дыхание! Артериальная кровь от легких через легочные вены попадает в левое предсердие, тогда как от кожи — через полые вены в венозный синус и затем в правое предсердие. В жизни земноводных нередки ситуации, когда этим животным приходится использовать в основном (или даже исключительно) кожное дыхание: так происходит при нырянии и длительном пребывании под водой. В этом случае легкие уже не снабжают организм кислородом, а сами его потребляют. Если бы желудочек сердца земноводных был разделен, артериальная кровь от кожи поступала бы по легочным артериям в легкие, а наиболее нуждающиеся в кислороде органы головы оказались бы его лишенными. Поэтому земноводным необходимо именно такое сердце, имеющее механизм для оптимального перераспределения потоков крови в соответствии с потребностями и физиологическим состоянием животных. При преимущественно легочном дыхании, напротив, целесообразно разделение крови, поступающей в желудочек сердца из разных предсердий. В полость желудочка от его стенок выступают мышечные гребни, которые частично разделяют ее на небольшие отсеки. Это препятствует смешиванию крови. В результате при сокращении желудочка в артериальный конус сначала поступает относительно наименее обогащенная кислородом кровь из правой части желудочка, затем смешанная и, наконец, наиболее богатая кислородом артериальная кровь из левой стороны желудочка. При легочном дыхании первая (относительно наиболее венозная) порция крови направляется в ближайшие к сердцу легочные (или кожно-легочные) артерии, вторая — в дуги аорты и третья (наиболее артериальная) — в сонные артерии. Если же животное находится под водой и дышит только кожей, несколько изменяется соотношение фаз пульсации желудочка и артериального конуса, в результате чего в легочные артерии подается относительно меньше крови, а в желудочке кровь из левого и правого предсердий сильнее смешивается, что позволяет, как было указано выше, оптимально использовать артериальную кровь, поступающую от кожи. У пресмыкающихся в связи с усовершенствованием механизма вентиляции легких (развитие мощного легочного насоса — грудной клетки) необходимость в кожном дыхании отпадает и появляется принципиальная возможность разделения артериального и венозного потоков крови. Однако у большинства рептилий перегородка в желудочке сердца остается неполной, и кровь там может смешиваться. Благодаря этой особенности у пресмыкающихся сохраняется возможность перераспределения крови в сердце и регуляции поступления больших или меньших ее количеств в разные сосуды. А это очень важно, и вот почему. Большое значение для пресмыкающихся имеет особая форма терморегуляции, называемая гелиотермией, — повышение температуры тела до оптимального уровня путем обогревания в лучах солнца (инсоляции). При инсоляции физиологически выгодно направлять больше крови в периферические сосуды (в частности, к кожным покровам) и меньше — в легкие! где испарение влаги приводит к некоторому понижению температуры тела. Напротив, когда животные, достигнув оптимальной температуры тела, переходят к активной деятельности (поиски пищи, перемещения по территории и т. п.), необходимо больше крови направлять к легким. В соответствии с этими потребностями и регулируется поток крови в малом и большом кругах кровообращения. Брюшная аорта у предков рептилий разделилась на три артериальных ствола, начинающихся от сердца: левую и правую дуги аорты (сливающиеся в спинную аорту) и общий ствол легочных артерий.
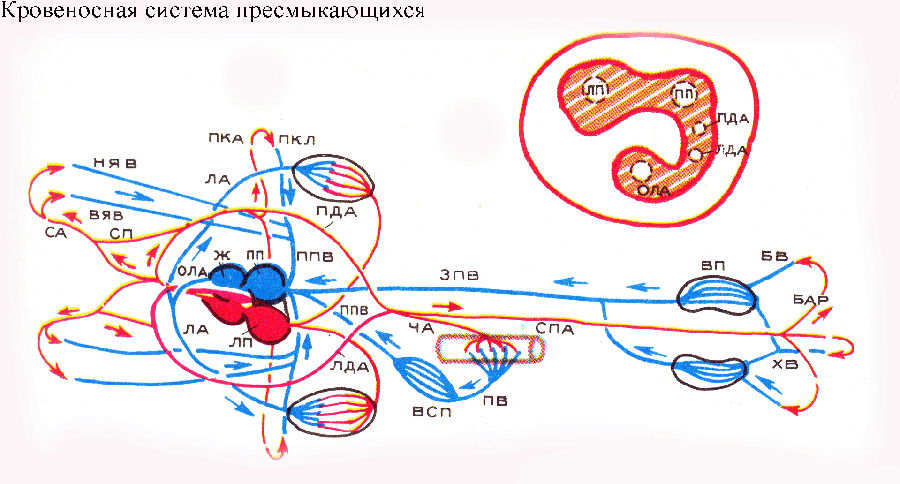
Правая дуга аорты получает из желудочка сердца наиболее чистую артериальную кровь; в связи с этим сонные и подключичные артерии (несущие кровь к голове и к передним конечностям) у рептилий начинаются от правой дуги аорты. Основные вены в принципе соответствуют таковым амфибий. Сердце у большинства пресмыкающихся трехкамерное: два предсердия и желудочек. Характерные для земноводных венозная пазуха и артериальный конус редуцировались. В левое предсердие впадают легочные вены, в правое — три полые вены. В желудочке имеется неполная перегородка довольно сложной формы, расположенная почти в горизонтальной плоскости и разделяющая желудочек на спинной и брюшной отделы. Оба предсердия впадают в спинной отдел желудочка. Распределение крови в желудочке происходит следующим образом. Первым сокращается правое предсердие, и венозная кровь из него стекает в брюшной отдел желудочка, от которого начинается общий ствол легочных артерий. Затем сокращается левое предсердие, артериальная кровь из которого заполняет спинной отдел желудочка. От этого отдела начинаются обе дуги аорты, причем начало левой дуги расположено правее и ниже начала правой (затем, по ходу, дуги аорты перекрещиваются). При систоле желудочка сердца его перегородка полностью отделяет брюшной отдел от спинного, так что в обе дуги аорты поступает артериальная кровь, а в легочные артерии — венозная. Если же при инсоляции требуется направить меньше крови в легочные артерии и больше — в дуги аорты, просвет общего ствола легочных артерий сужается (сокращением гладкой мускулатуры в его стенках) и часть венозной крови из брюшного отдела желудочка возвращается в спинной. В левую дугу аорты в этом случае попадает смешанная кровь. Как было уже указано, голова и передние конечности всегда получают артериальную кровь через правую дугу аорты. Нетрудно заметить, что полное анатомическое разделение желудочка сердца такой горизонтальной перегородкой невозможно, так как при этом легочные артерии оказались бы изолированы от потока крови. Эту проблему решили высшие рептилии — крокодилы: у их предков первоначальная горизонтальная перегородка в желудочке сердца отчасти редуцировалась, отчасти преобразовалась в новую, вертикальную перегородку, полностью разделившую желудочек на левую и правую камеры. Причем от левого желудочка начинается правая дуга аорты, а от правого — левая дуга аорты и общий ствол легочных артерий. Артериальные сосуды, начинающиеся от правого желудочка, получают из сердца венозную кровь. Однако когда крокодилы находятся на поверхности воды или на суше и нормально дышат, по левой дуге аорты венозная кровь проходит только до перекреста двух дуг аорты, где между последними имеется связь — так называемое Паниццево отверстие. Благодаря более высокому давлению в правой дуге аорты артериальная кровь из нее через это отверстие поступает и в левую дугу аорты. А при нырянии и при инсоляции у крокодилов происходит перераспределение крови в магистральных артериях, как и у других пресмыкающихся: сужается просвет легочных артерий, повышается давление крови в левом желудочке и левой дуге аорты и венозная кровь начинает поступать по левой дуге дальше Паниццева отверстия. Отметим, что левая дуга у крокодилов не сливается с правой, а продолжается в чревную артерию, снабжающую кровью кишечник. Спинную же аорту образует лишь правая дуга. Предки птиц в филогенезе древних рептилий обособились от группы псевдозухий, от которых возникли и крокодилы. Не случайно поэтому кровеносная система птиц обнаруживает определенное сходство с таковой крокодилов, как бы продолжая намеченное у последних направление эволюционных перестроек. У птиц, как и у крокодилов, сердце четырехкамерное, и желудочек разделен полной вертикальной перегородкой на левую и правую камеры.
Птицы приобрели гомойотермию — способность поддерживать температуру своего тела на постоянном уровне «теплокровность». Поэтому для них отпала необходимость в инсоляции и перераспределении крови в сердце и магистральных артериях, столь важном для пресмыкающихся. Левая дуга аорты в этих условиях стала ненужной и полностью редуцировалась. От сердца птицы отходят два артериальных ствола:
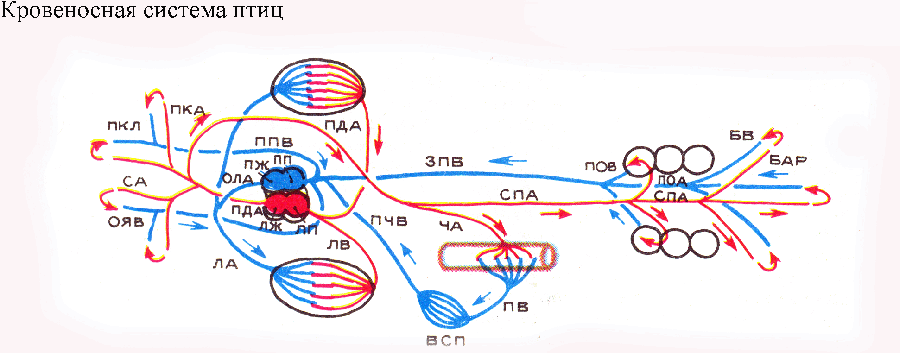
правая дуга аорты — от левого желудочка (получает артериальную кровь) и общий ствол легочных артерий — от правого желудочка (получает венозную кровь). Общая схема кровеносной системы птиц, таким образом, упрощена по сравнению с таковой рептилий. Морфо-физиологический прогресс здесь достигнут не усложнением, а упрощением и рационализацией системы. Воротная система почек у птиц подверглась редукции, и кровоснабжение почек осуществляется почечными артериями, отходящими от спинной аорты.
У предков млекопитающих брюшная аорта разделилась не на три, как у типичных пресмыкающихся, а на два артериальных ствола: общий ствол дуг аорты и общий ствол легочных артерий.
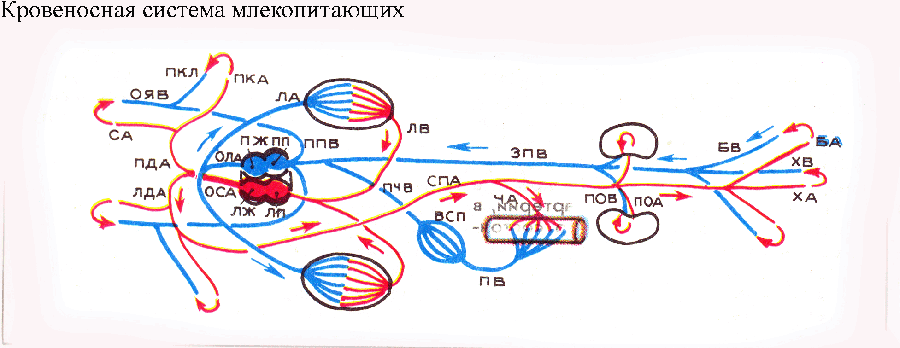
Это упростило задачу разделения крови в сердце млекопитающих, которое стало четырехкамерным, как у птиц. Желудочек сердца разделился вертикальной перегородкой на левый (от которого начинается общий ствол дуг аорты) и правый (откуда берет начало легочный ствол). Впереди общий ствол дуг аорты разделяется на левую дугу аорты (продолжающуюся в спинную аорту) и безымянную артерию, которая гомологична редуцированной правой дуге аорты — она не достигает спинной аорты и переходит в правую подключичную артерию. Сонные артерии начинаются симметрично: левая — от левой дуги аорты, правая — от безымянной артерии (у некоторых видов млекопитающих обе сонные артерии начинаются от общего ствола с правой подключичной артерией, который называется в этом случае плечеголовным). В венозной системе млекопитающих, как и у птиц, редуцировалась воротная система почек. Возможно, в обоих случаях это связано с повышением кровяного давления (по сравнению с низшими позвоночными). При видимом сходстве кровеносных систем млекопитающих и птиц (четырехкамерное сердце, единственная дуга аорты образует спинную аорту, редукция воротной системы почек) между ними имеются глубокие различия. У птиц от левого желудочка сердца начинается правая дуга аорты, а левая утрачена, тогда как у млекопитающих от левого желудочка начинается общий ствол дуг аорты, левая дуга развита полностью, а правая редуцирована. Эти различия связаны с длительной независимой эволюцией предков птиц и млекопитающих, в ходе которой сходные прогрессивные черты кровеносной системы были приобретены разными путями. Вероятно, у общих предков птиц и млекопитающих еще сохранялся единый желудочек сердца и артериальный конус, а также короткий общий артериальный ствол — брюшная аорта. Разделение желудочка сердца на камеры и брюшной аорты на магистральные артерии происходило самостоятельно и по-разному в филогенетических стволах предковых форм рептилий, ведущих к птицам и млекопитающим. Развитие горизонтальной внутрижелудочковой перегородки в сердце типичных пресмыкающихся интересно как пример инадаптивного пути эволюционных преобразований. По В. О. Ковалевскому, при инадаптивном развитии формирующееся приспособление несет в себе предпосылки для возникновения каких-либо внутренних конструктивных дефектов, обнаруживающихся не сразу, но постепенно нарастающих по мере развития. Инадаптивная эволюция является результатом своего рода «неудачного» выбора направления эволюционных изменений. А такой «неудачный выбор» вполне вероятен:
естественный отбор благоприятствует любым изменениям, которые дают какие-либо преимущества организмам в борьбе за существование в данный исторический момент, но не может «предвидеть» ни результатов дальнейшей эволюции в том или другом направлении, ни новых форм взаимодействия организма с внешней средой, которые сложатся в будущем. На уровне организации пресмыкающихся их неполная внутрижелудочковая перегородка в целом отвечает физиологическим потребностям этих животных, разделяя артериальную и венозную кровь и в то же время позволяя перераспределять потоки крови в соответствии с характером активности. Однако из-за своего положения относительно устий предсердий и мест отхождения артерий эта перегородка не может полностью завершить разделение желудочка сердца на артериальную и венозную камеры. На базе такого устройства сердца его дальнейшая прогрессивная эволюция затруднена и требует довольно сложных перестроек (произошедших у крокодилов и птиц). В то же время на первых этапах возникновения внутрижелудочковой перегородки ее вертикальное или горизонтальное положение, так же как и разделение брюшной аорты на три или на два магистральных ствола, вероятно, не имели существенных преимуществ друг перед другом. Однако путь эволюционных преобразований сердца у предков млекопитающих был более перспективным для полного разделения желудочка, что обнаружилось на более поздних этапах эволюции. В эволюции позвоночных, как отмечалось выше, в целом происходило общее усложнение и совершенствование кровеносной системы. Однако не все эволюционные перестройки кровеносной системы представляют собой ароморфозы (т. е. изменения на пути морфо-физиологического прогресса, по А. Н. Северцову). Усложнение организации само по себе еще не является прогрессивным изменением. Иногда оно служит своего рода компенсацией за несовершенство какой-либо функции или представляет собой результат развития частного приспособления (идиоадаптации, по А. Н. Северцову) к специфическому образу жизни. Именно так следует оценивать появление второго (малого) круга кровообращения и усложнение строения сердца у двоякодышащих рыб. Лишь в тех случаях, когда усложнение строения связано с интенсификацией функций, с повышением уровня жизнедеятельности (выражающимся в интенсификации процессов обмена веществ) и возрастанием степени гомеостаза (независимости внутренней среды организма от внешних условий), можно говорить об ароморфозах.
В эволюции кровеносной системы хордовых несомненными ароморфозами были развитие сердца у древнейших позвоночных, интенсификация его работы и формирование замкнутости кровеносной системы у челюстноротых, преобразования сердца и магистральных сосудов при разделении потоков артериальной и венозной крови у пресмыкающихся, птиц и млекопитающих.
По статье Н.Н. Иорданского — доктора биологических наук
←На предыдущую страницу
Эволюция органов кровообращения
Кровообращение хордовых
Цели:
- Определить, что такое кровообращение?
- Определить, как меняются органы кровообращения с изменением среды обитания?
- У простейших перенос веществ осуществляется за счет диффузии молекул и движения цитоплазмы.
- У низших многоклеточных животных, имеющих относительно простое строение (губки, кишечнополостные, плоские черви), перенос молекул также обеспечивается диффузией.
- У животных, ведущих активный образ жизни, имеющих сложное строение и интенсивный обмен веществ, в ходе эволюции сформировались высокоэффективные специальные системы внутреннего транспорта веществ — кровеносная и лимфатическая.
В ходе эволюции многоклеточных животных кровеносная система сформировалась на месте рудиментов первичной полости тела, вытесненной у высших животных вторичной полостью тела, или целомом. Кровеносная система приняла на себя в организме высших животных универсальную транспортную роль передвижение питательных веществ, усвоенных органами пищеварения, и кислорода от органов дыхания ко всем органам тела, перенос продуктов метаболизма к органам выделения, углекислоты — к органам дыхания, доставка гормонов от эндокринных желез к органам, где данные гормоны будут действовать. В силу этого кровеносная система стала и одной из важнейших интегрирующих систем организма, обеспечивающих его целостность. Кровеносная система имеется у немертин кольчатых червей, моллюсков, членистоногих. иглокожих, полухордовых и хордовых. Ее строение у животных разных типов сильно варьирует. Два основных продольных сосуда — спинной и брюшной, тянущиеся соответственно над и под кишечником, связаны друг с другом поперечными сосудами; последние кольцеобразно охватывают кишечник и дают ответвления к покровам и различным внутренним органам. У животных всех типов, условно объединяемых названием «беспозвоночные» кровь по брюшному сосуду течет назад, а по спинному вперед. У хордовых наоборот: по брюшному сосуду кровь течет вперед, а по спинному назад. Пульсирующей функцией, обеспечивающей движение крови, у хордовых обладает брюшной сосуд или его производное — сердце, у беспозвоночных эту функцию несет спинной сосуд. У общих предков вторичноротых животных (к ним принадлежат полухордовые, иглокожие и хордовые) кровеносная система имела очень примитивное строение, представляя собой систему лакун, т. е. полостей без эпителизированных стенок, где направление кровотока еще не установилось. В дальнейшем совершенствование кровеносной системы происходило независимо и по-разному в различных ветвях ствола вторичноротых. Поэтому состояние кровеносной системы хордовых сформировалось особым путем и не может быть выведено из развитой кровеносной системы беспозвоночных — как вторичноротых, так и (тем более) первичноротых, к которым принадлежат кольчатые черви, членистоногие, моллюски и др.
|
артериальная кровь, синим цветом — венозная; |
|
У типичного представителя бесчерепных — ланцетника — под глоткой тянется продольный сосуд — брюшная аорта, играющая роль основного пульсирующего органа. По брюшной аорте кровь течет вперед. От этого магистрального сосуда начинаются многочисленные (около 100 пар) жаберные артерии, которые расположены в перегородках между жаберными щелями, пронизывающими боковые стенки глотки. Основания жаберных артерий пульсируют, давая добавочный импульс току крови. Через жаберные щели непрерывно процеживается вода, и здесь происходит газообмен между кровью и водой: кровь в жаберных артериях обогащается кислородом и отдает углекислый газ, т. е. становится артериальной. Обогащенную кислородом кровь жаберные артерии доставляют на спинную сторону глотки, где они впадают в пару продольных сосудов — корни спинной аорты. Впереди корни аорты переходят в сонные артерии, несущие арте кровь в головной конец тела, корни аорты впадают в продольный спинной сосуд — спинную аорту, по которой артериальная кровь течет назад. От спинной аорты отходят артерии к различным внутренним органам и к кожным покровам, где артерии распадаются на капилляры. Из хвостовой области тела кровь собирается в хвостовую вену, впереди впадающую в подкишечную вену. Последняя собирает кровь от кишечника, а затем в печеночном выросте кишки вновь распадается на капилляры, образующие воротную систему печени. Из нее печеночная вена несет кровь в брюшную аорту. У ланцетника имеются еще две пары магистральных вен: передние и задние кардинальные, тянущиеся от переднего и заднего концов тела к средней его части, где они впадают в пару коротких Кювьеровых протоков. Последние несут кровь в задний конец брюшной аорты. Таким образом, брюшная аорта получает обедненную кислородом (венозную) кровь.
У второго подтипа хордовых — оболочников — кровеносная система подверглась редукции. Как и целый ряд других органов, в связи с переходом к неподвижному (прикрепленному) образу жизни на морском дне. Освоение такого образа жизни в эволюции обычно ведет к морфо-физиологическому регрессу. У типичных представителей оболочников асцидий — под желудком располагается трубкообразное сердце, от которого отходят два магистральных сосуда: спереди — жаберная артерия, тянущаяся по брюшной стороне глотки и отдающая ветки к перегородкам между жаберными отверстиями, позади — кишечная артерия, которая разветвляется на сосуды, идущие к внутренним органам. В конечном итоге артерии впадают в лакуны, т. е. кровеносная система оболочников не замкнута. Одна из таких лакун — жаберно-кишечный синус — лежит над глоткой и собирает кровь из жаберной области. Интересно, что направление пульсации сердца у оболочников периодически меняется: сердце попеременно гонит кровь то вперед, в жаберную артерию, то назад, в кишечную. Некоторые ученые видят в этом примитивную черту — еще не установившееся направление кровотока. Уникальной особенностью оболочников является их кровяной пигмент — гемованадин, в котором вместо ионов железа (как в гемоглобине остальных хордовых) или меди (как в гемоцианине беспозвоночных ряда групп) используются ионы ванадия. Эволюция третьего подтипа хордовых — позвоночных — в целом шла в направлении морфо-физиологического прогресса, что, в частности, ярко проявилось в последовательном совершенствовании кровеносной системы: ее дифференциации, интеграции, интенсификации функций в ходе филогенеза позвоночных. У всех позвоночных центральным пропульсаторным органом является сердце. У миног, представителей наиболее примитивного класса современных позвоночных — круглоротых, сердце имеет три отдела (камеры):
венозный синус (венозная пазуха), предсердие и желудочек . От желудочка отходит брюшная аорта, начало которой расширено (луковица аорты). Впереди брюшная аорта разделяется на пару поджаберных артерий. От брюшной аорты и поджаберных артерий начинаются 8 пар приносящих жаберных артерий, распадающихся в жабрах на капилляры (этого не было у бесчерепных). От жабер обогащенная кислородом кровь поступает в выносящие жаберные артерии, которые впадают в спинную аорту. В целом схема кровеносной системы круглоротых сходна с таковой бесчерепных, но отличается наличием еще одной магистральной вены (нижней яремной), выносящей кровь из вентральной части головы, и отсутствием Кювьеровых протоков (специфическая черта круглоротых). Обе пары кардинальных вен, печеночная и нижняя яремная вены впадают непосредственно в венозную пазуху. Еще одной специфической особенностью кровеносной системы круглоротых, является ее неполная замкнутость: жаберные мешки окружены околожаберными синусами, заполненными кровью и представляющими собой кровеносные лакуны. В связи с этим активное участие в циркуляции крови принимают мышцы жаберного аппарата, сокращение которых выталкивает кровь из лакун и сосудов жаберной области. У другой группы круглоротых — миксин — работе сердца помогают также пульсирующие участки ряда вен (кардинальных, хвостовой и воротной вены печени).
У всех остальных позвоночных — челюсторотых — по сравнению с круглоротыми был достигнут существенный прогресс в организации кровеносной системы. Она стала полностью замкнутой, сердце полностью приняло на себя роль центрального пульсирующего органа. Это сделало возможным повышение кровяного давления и интенсификацию кровотока, что способствовало общему повышению уровня жизнедеятельности. У хрящевых рыб (акулы, скаты, химеры) сердце состоит из 4 камер:
к З камерам, известным нам по круглоротым, добавился еще артериальный конус. Функция его — распределение крови между передними и задними приносящими жаберными артериями. Парные поджаберные сосуды, имевшиеся у круглоротых, редуцированы. У большинства хрящевых рыб жабры сохраняются лишь на 4 жаберных дугах и на подъязычной дуге висцерального аппарата; соответственно, имеется 5 пар приносящих жаберных артерий. Как и у бесчерепных, магистральные вены впадают не прямо в сердце, а в пару Кювьеровых протоков. В отличие от круглоротых, у хрящевых рыб нижняя яремная вена парная, а на пути крови из хвостовой области имеется еще одна капиллярная сеть — в почках (воротная система почек).
Класс костных рыб объединяет ряд филогенетически рано обособившихся друг от друга групп, у которых кровеносные системы существенно различаются по строению. У наиболее распространенных и много численных костных рыб (к ним относится большинство наших обычных пресноводных рыб подвергся редукции артериальный конус сердца, и оно стало трехкамерным, как у круглоротых. В начале брюшной аорты имеется расширение (луковица аорты). Развиты лишь 4 пары приносящих жаберных артерий. Воротная система левой почки редуцирована. Наземные позвоночные возникли от древних кистеперых рыб, обитавших в пресных мелководных, богатых растительностью тропических водоемах, отчасти заболоченных. В таких водоемах часто возникал дефицит растворенного в воде кислорода (в связи с усиленным гниением отмиравших растений). Важным приспособлением к жизни в этих условиях было развитие легких, позволявших при дефиците кислорода в воде дышать атмосферным воздухом. Среди современных костных рыб близкий образ жизни ведут родственные кистеперым рыбам двоякодышащие, тогда как единственный доживший до наших дней вид кистеперых — латимерия — обитает в морских глубинах и легкие для дыхания не использует. Морфо-физиологической особенностью рыб, дышащих и жабрами, и легкими (древние кистеперые и современные двоякодышащие), является решение проблемы рационального использования обоих органов газообмена. Дело в том, что при нормальном для рыб жаберном дыхании обогащенная кислородом кровь из передних жабер поступает в голову, тогда как из задних — в спинную аорту, несущую кровь назад по всему телу. Легкие сформировались позади глотки, т. е. позади самых задних жабер. И поэтому если кровь из легких, как из жабер, направить прямо в корни аорты, то эта артериальная кровь поступит в спинную аорту, а органы головы (и, в частности, головной мозг, особенно н уждающийся в постоянном поступлении кислорода) не смогут ее использовать. Поэтому возникает необходимость нового перераспределения крови. Эта проблема была решена возвращением артериальной крови из легких в сердце (т. е. образованием дополнительного легочного, или малого, круга кровообращения) и формированием в сердце особых приспособлений для перераспределения крови.
У двоякодышащих
сердце состоит из венозного синуса (получающего, как обычно у рыб, венозную кровь), предсердия, желудочка и артериального конуса. Венозный синус впадает в правую половину предсердия, тогда как в левую его половину впадают легочные вены, приносящие артериальную кровь из легких. В предсердии, желудочке и артериальном конусе имеются неполные внутренние перегородки (в частности, спиральный клапан артериального конуса), позволяющие разделить кровь, поступившую из левой и правой половин предсердия. Из левой половины предсердия артериальная кровь поступает в нижнюю часть сильно укороченной брюшной аорты и по ней — к двум передним парам приносящих жаберных артерий; затем эта кровь поступает по сонным артериям в голову. Из правой половины предсердия венозная кровь направляется в верхнюю часть брюшной аорты и по ней — к двум задним (ближайшим к сердцу) парам жаберных артерий. Подчеркнем, что легочные артерии начинаются от четвертой (самой задней) пары выносящих жаберных артерий. Механизм распределения крови в сердце двоякодышащих рыб лабилен и позволяет направлять больше или меньше крови в передние или задние жаберные артерии в зависимости от физиологического состояния рыбы и от того, какой орган (легкие или жабры) в данный момент работают интенсивнее. В венозной системе у двоякодышащих рыб редуцирована правая задняя полая вена, зато появился новый магистральный венозный ствол — задняя полая вена, несущая кровь из воротной системы почек прямо в венозный синус, минуя Кювьеровы протоки. У наземных позвоночных (тетрапод) произошли значительные изменения в строении кровеносной системы по сравнению с рыбами (хотя общее направление этих перестроек намечено уже у двоякодышащих). У всех тетрапод брюшная аорта отчасти редуцирована, отчасти разделена на несколько обособленных артериальных стволов, начинающихся от сердца, причем это разделение произошло по-разному у предков современных земноводных, пресмыкающихся и млекопитающих. У современных земноводных
от артериального конуса сердца начинаются три пары сосудов: сонные артерии, дуги аорты (которые на спинной стороне сливаются, образуя спинную аорту) и легочные (а у бесхвостых амфибий — кожно-легочные) артерии. Первые из них соответствуют 1 паре жаберных артерий с собственно сонными артериями рыб; вторые — II паре жаберных артерий с корнями аорты рыб; третьи — IV паре жаберных артерий с легочными артериями рыб (у хвостатых амфибий иногда сохраняются две пары дуг аорты, соответствующие II и III парам жаберных артерий). У рыб все эти сосуды связаны друг с другом над глоткой корнями аорты; такая связь сохраняется и у некоторых тетрапод в виде сонных протоков между сонными артериями и дугами аорты, а также Боталловых протоков между дугами аорты и легочными артериями. По сонным артериям кровь направляется к голове; по дугам аорты — в спинную аорту и по ней к различным органам тела; легочные (или кожно-легочные) артерии обеспечивают кровоснабжение органов дыхания. У земноводных несовершенство механизма вентиляции легких не позволяет этим животным ограничиться легочным дыханием, и кожа у них является важным дополнительным органом дыхания. У хвостатых и безногих амфибий, легочные артерии не объединены с кожными. Кровоснабжение кожных покровов, как и у других позвоночных, осуществляется ветвями спинной аорты.
Венозная система земноводных отличается от таковой рыб редукцией задних кардинальных вен, которые замещаются задней полой веной, получающей кровь из воротных систем почек и печени и доставляющей ее в венозную пазуху сердца. Передние кардинальные вены у наземных позвоночных сохраняются, но получают новое название: их традиционно именуют верхними яремными венами. Кювьеровы протоки у тетрапод называют передними полыми венами. Сердце земноводных состоит из пяти отделов: венозного синуса (пазухи), левого и правого предсердий, желудочка и артериального конуса. В венозный синус впадают передние и задняя полые вены. В левое предсердие впадают легочные вены, приносящие артериальную кровь из легких. В желудочке кровь из левого и правого предсердий в определенной степени смешивается. Нередко это упрощенно объясняют примитивностью и несовершенством сердца земноводных, но в действительности дело обстоит сложнее и интереснее. Объединение в желудочке сердца земноводных крови, поступающей из левого и правого предсердий, имеет важное функциональное значение: если бы такого смешивания крови не было, земноводным не удалось бы эффективно использовать кожное дыхание! Артериальная кровь от легких через легочные вены попадает в левое предсердие, тогда как от кожи — через полые вены в венозный синус и затем в правое предсердие. В жизни земноводных нередки ситуации, когда этим животным приходится использовать в основном (или даже исключительно) кожное дыхание: так происходит при нырянии и длительном пребывании под водой. В этом случае легкие уже не снабжают организм кислородом, а сами его потребляют. Если бы желудочек сердца земноводных был разделен, артериальная кровь от кожи поступала бы по легочным артериям в легкие, а наиболее нуждающиеся в кислороде органы головы оказались бы его лишенными. Поэтому земноводным необходимо именно такое сердце, имеющее механизм для оптимального перераспределения потоков крови в соответствии с потребностями и физиологическим состоянием животных. При преимущественно легочном дыхании, напротив, целесообразно разделение крови, поступающей в желудочек сердца из разных предсердий. В полость желудочка от его стенок выступают мышечные гребни, которые частично разделяют ее на небольшие отсеки. Это препятствует смешиванию крови. В результате при сокращении желудочка в артериальный конус сначала поступает относительно наименее обогащенная кислородом кровь из правой части желудочка, затем смешанная и, наконец, наиболее богатая кислородом артериальная кровь из левой стороны желудочка. При легочном дыхании первая (относительно наиболее венозная) порция крови направляется в ближайшие к сердцу легочные (или кожно-легочные) артерии, вторая — в дуги аорты и третья (наиболее артериальная) — в сонные артерии. Если же животное находится под водой и дышит только кожей, несколько изменяется соотношение фаз пульсации желудочка и артериального конуса, в результате чего в легочные артерии подается относительно меньше крови, а в желудочке кровь из левого и правого предсердий сильнее смешивается, что позволяет, как было указано выше, оптимально использовать артериальную кровь, поступающую от кожи. У пресмыкающихся в связи с усовершенствованием механизма вентиляции легких (развитие мощного легочного насоса — грудной клетки) необходимость в кожном дыхании отпадает и появляется принципиальная возможность разделения артериального и венозного потоков крови. Однако у большинства рептилий перегородка в желудочке сердца остается неполной, и кровь там может смешиваться. Благодаря этой особенности у пресмыкающихся сохраняется возможность перераспределения крови в сердце и регуляции поступления больших или меньших ее количеств в разные сосуды. А это очень важно, и вот почему. Большое значение для пресмыкающихся имеет особая форма терморегуляции, называемая гелиотермией, — повышение температуры тела до оптимального уровня путем обогревания в лучах солнца (инсоляции). При инсоляции физиологически выгодно направлять больше крови в периферические сосуды (в частности, к кожным покровам) и меньше — в легкие! где испарение влаги приводит к некоторому понижению температуры тела. Напротив, когда животные, достигнув оптимальной температуры тела, переходят к активной деятельности (поиски пищи, перемещения по территории и т. п.), необходимо больше крови направлять к легким. В соответствии с этими потребностями и регулируется поток крови в малом и большом кругах кровообращения. Брюшная аорта у предков рептилий разделилась на три артериальных ствола, начинающихся от сердца: левую и правую дуги аорты (сливающиеся в спинную аорту) и общий ствол легочных артерий.
Правая дуга аорты получает из желудочка сердца наиболее чистую артериальную кровь; в связи с этим сонные и подключичные артерии (несущие кровь к голове и к передним конечностям) у рептилий начинаются от правой дуги аорты. Основные вены в принципе соответствуют таковым амфибий. Сердце у большинства пресмыкающихся трехкамерное: два предсердия и желудочек. Характерные для земноводных венозная пазуха и артериальный конус редуцировались. В левое предсердие впадают легочные вены, в правое — три полые вены. В желудочке имеется неполная перегородка довольно сложной формы, расположенная почти в горизонтальной плоскости и разделяющая желудочек на спинной и брюшной отделы. Оба предсердия впадают в спинной отдел желудочка. Распределение крови в желудочке происходит следующим образом. Первым сокращается правое предсердие, и венозная кровь из него стекает в брюшной отдел желудочка, от которого начинается общий ствол легочных артерий. Затем сокращается левое предсердие, артериальная кровь из которого заполняет спинной отдел желудочка. От этого отдела начинаются обе дуги аорты, причем начало левой дуги расположено правее и ниже начала правой (затем, по ходу, дуги аорты перекрещиваются). При систоле желудочка сердца его перегородка полностью отделяет брюшной отдел от спинного, так что в обе дуги аорты поступает артериальная кровь, а в легочные артерии — венозная. Если же при инсоляции требуется направить меньше крови в легочные артерии и больше — в дуги аорты, просвет общего ствола легочных артерий сужается (сокращением гладкой мускулатуры в его стенках) и часть венозной крови из брюшного отдела желудочка возвращается в спинной. В левую дугу аорты в этом случае попадает смешанная кровь. Как было уже указано, голова и передние конечности всегда получают артериальную кровь через правую дугу аорты. Нетрудно заметить, что полное анатомическое разделение желудочка сердца такой горизонтальной перегородкой невозможно, так как при этом легочные артерии оказались бы изолированы от потока крови. Эту проблему решили высшие рептилии — крокодилы: у их предков первоначальная горизонтальная перегородка в желудочке сердца отчасти редуцировалась, отчасти преобразовалась в новую, вертикальную перегородку, полностью разделившую желудочек на левую и правую камеры. Причем от левого желудочка начинается правая дуга аорты, а от правого — левая дуга аорты и общий ствол легочных артерий. Артериальные сосуды, начинающиеся от правого желудочка, получают из сердца венозную кровь. Однако когда крокодилы находятся на поверхности воды или на суше и нормально дышат, по левой дуге аорты венозная кровь проходит только до перекреста двух дуг аорты, где между последними имеется связь — так называемое Паниццево отверстие. Благодаря более высокому давлению в правой дуге аорты артериальная кровь из нее через это отверстие поступает и в левую дугу аорты. А при нырянии и при инсоляции у крокодилов происходит перераспределение крови в магистральных артериях, как и у других пресмыкающихся: сужается просвет легочных артерий, повышается давление крови в левом желудочке и левой дуге аорты и венозная кровь начинает поступать по левой дуге дальше Паниццева отверстия. Отметим, что левая дуга у крокодилов не сливается с правой, а продолжается в чревную артерию, снабжающую кровью кишечник. Спинную же аорту образует лишь правая дуга. Предки птиц в филогенезе древних рептилий обособились от группы псевдозухий, от которых возникли и крокодилы. Не случайно поэтому кровеносная система птиц обнаруживает определенное сходство с таковой крокодилов, как бы продолжая намеченное у последних направление эволюционных перестроек. У птиц, как и у крокодилов, сердце четырехкамерное, и желудочек разделен полной вертикальной перегородкой на левую и правую камеры.
Птицы приобрели гомойотермию — способность поддерживать температуру своего тела на постоянном уровне «теплокровность». Поэтому для них отпала необходимость в инсоляции и перераспределении крови в сердце и магистральных артериях, столь важном для пресмыкающихся. Левая дуга аорты в этих условиях стала ненужной и полностью редуцировалась. От сердца птицы отходят два артериальных ствола:
правая дуга аорты — от левого желудочка (получает артериальную кровь) и общий ствол легочных артерий — от правого желудочка (получает венозную кровь). Общая схема кровеносной системы птиц, таким образом, упрощена по сравнению с таковой рептилий. Морфо-физиологический прогресс здесь достигнут не усложнением, а упрощением и рационализацией системы. Воротная система почек у птиц подверглась редукции, и кровоснабжение почек осуществляется почечными артериями, отходящими от спинной аорты.
У предков млекопитающих брюшная аорта разделилась не на три, как у типичных пресмыкающихся, а на два артериальных ствола: общий ствол дуг аорты и общий ствол легочных артерий.
Это упростило задачу разделения крови в сердце млекопитающих, которое стало четырехкамерным, как у птиц. Желудочек сердца разделился вертикальной перегородкой на левый (от которого начинается общий ствол дуг аорты) и правый (откуда берет начало легочный ствол). Впереди общий ствол дуг аорты разделяется на левую дугу аорты (продолжающуюся в спинную аорту) и безымянную артерию, которая гомологична редуцированной правой дуге аорты — она не достигает спинной аорты и переходит в правую подключичную артерию. Сонные артерии начинаются симметрично: левая — от левой дуги аорты, правая — от безымянной артерии (у некоторых видов млекопитающих обе сонные артерии начинаются от общего ствола с правой подключичной артерией, который называется в этом случае плечеголовным). В венозной системе млекопитающих, как и у птиц, редуцировалась воротная система почек. Возможно, в обоих случаях это связано с повышением кровяного давления (по сравнению с низшими позвоночными). При видимом сходстве кровеносных систем млекопитающих и птиц (четырехкамерное сердце, единственная дуга аорты образует спинную аорту, редукция воротной системы почек) между ними имеются глубокие различия. У птиц от левого желудочка сердца начинается правая дуга аорты, а левая утрачена, тогда как у млекопитающих от левого желудочка начинается общий ствол дуг аорты, левая дуга развита полностью, а правая редуцирована. Эти различия связаны с длительной независимой эволюцией предков птиц и млекопитающих, в ходе которой сходные прогрессивные черты кровеносной системы были приобретены разными путями. Вероятно, у общих предков птиц и млекопитающих еще сохранялся единый желудочек сердца и артериальный конус, а также короткий общий артериальный ствол — брюшная аорта. Разделение желудочка сердца на камеры и брюшной аорты на магистральные артерии происходило самостоятельно и по-разному в филогенетических стволах предковых форм рептилий, ведущих к птицам и млекопитающим. Развитие горизонтальной внутрижелудочковой перегородки в сердце типичных пресмыкающихся интересно как пример инадаптивного пути эволюционных преобразований. По В. О. Ковалевскому, при инадаптивном развитии формирующееся приспособление несет в себе предпосылки для возникновения каких-либо внутренних конструктивных дефектов, обнаруживающихся не сразу, но постепенно нарастающих по мере развития. Инадаптивная эволюция является результатом своего рода «неудачного» выбора направления эволюционных изменений. А такой «неудачный выбор» вполне вероятен:
естественный отбор благоприятствует любым изменениям, которые дают какие-либо преимущества организмам в борьбе за существование в данный исторический момент, но не может «предвидеть» ни результатов дальнейшей эволюции в том или другом направлении, ни новых форм взаимодействия организма с внешней средой, которые сложатся в будущем. На уровне организации пресмыкающихся их неполная внутрижелудочковая перегородка в целом отвечает физиологическим потребностям этих животных, разделяя артериальную и венозную кровь и в то же время позволяя перераспределять потоки крови в соответствии с характером активности. Однако из-за своего положения относительно устий предсердий и мест отхождения артерий эта перегородка не может полностью завершить разделение желудочка сердца на артериальную и венозную камеры. На базе такого устройства сердца его дальнейшая прогрессивная эволюция затруднена и требует довольно сложных перестроек (произошедших у крокодилов и птиц). В то же время на первых этапах возникновения внутрижелудочковой перегородки ее вертикальное или горизонтальное положение, так же как и разделение брюшной аорты на три или на два магистральных ствола, вероятно, не имели существенных преимуществ друг перед другом. Однако путь эволюционных преобразований сердца у предков млекопитающих был более перспективным для полного разделения желудочка, что обнаружилось на более поздних этапах эволюции. В эволюции позвоночных, как отмечалось выше, в целом происходило общее усложнение и совершенствование кровеносной системы. Однако не все эволюционные перестройки кровеносной системы представляют собой ароморфозы (т. е. изменения на пути морфо-физиологического прогресса, по А. Н. Северцову). Усложнение организации само по себе еще не является прогрессивным изменением. Иногда оно служит своего рода компенсацией за несовершенство какой-либо функции или представляет собой результат развития частного приспособления (идиоадаптации, по А. Н. Северцову) к специфическому образу жизни. Именно так следует оценивать появление второго (малого) круга кровообращения и усложнение строения сердца у двоякодышащих рыб. Лишь в тех случаях, когда усложнение строения связано с интенсификацией функций, с повышением уровня жизнедеятельности (выражающимся в интенсификации процессов обмена веществ) и возрастанием степени гомеостаза (независимости внутренней среды организма от внешних условий), можно говорить об ароморфозах.
В эволюции кровеносной системы хордовых несомненными ароморфозами были развитие сердца у древнейших позвоночных, интенсификация его работы и формирование замкнутости кровеносной системы у челюстноротых, преобразования сердца и магистральных сосудов при разделении потоков артериальной и венозной крови у пресмыкающихся, птиц и млекопитающих.
По статье Н.Н. Иорданского — доктора биологических наук
←На предыдущую страницу